Mixed turf of bryozoans and erect sponges with Cylista elegans on tide-swept circalittoral rock
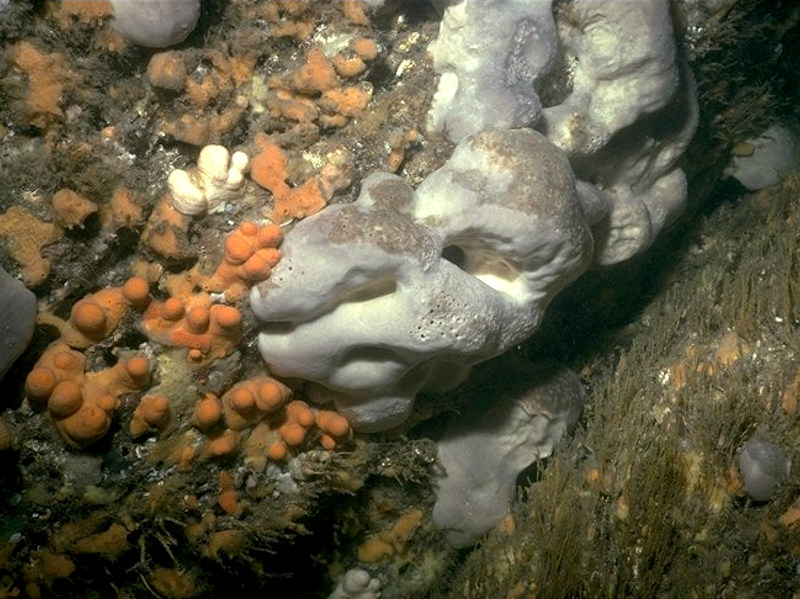
Mixed turf of bryozoans and erect sponges with Cylista elegans on tide-swept ciraclittoral rock
Photographer: Rohan Holt Copyright: Joint Nature Conservation Committee (JNCC)

| Researched by | John Readman, Kelsey Lloyd & Amy Watson | Refereed by | This information is not refereed |
|---|
Summary
UK and Ireland classification
Description
This variant is typically found on wave-exposed circalittoral bedrock and boulders, on steep slopes and upper faces in moderate tidal streams. This species-rich biotope is characterized by a dense sponge, hydroid and bryozoan turf and frequent Alcyonium digitatum. There are frequently large growths of Cliona celata and Pachymatisma johnstonia. Other species present in this diverse sponge community include Polymastia boletiformis, Haliclona viscosa, Polymastia mamilliaris, Scypha ciliata, Hemimycale columella and Dysidea fragilis. Axinellid sponges such as Stelligera stuposa and Raspailia ramosa may be present in low abundance, and are usually more abundant in deeper water. A dense hydroid turf forms a significant part of this biotope, with tufts of large hydroids such as Nemertesia antennina and Nemertesia ramosa frequently recorded. Other hydroid turf component species include Halecium halecinum, Aglaophenia tubulifera and Abietinaria abietina. Anemones are also well represented, with species such as Urticina felina, Cylista elegans and Metridium senile recorded. The cup coral Caryophyllia smithii and the anemone Corynactis viridis are also frequently seen. The bryozoan turf is composed predominantly of Alcyonidium diaphanum and Flustra foliacea, whilst crustose species such as Parasmittina trispinosa contribute to a lesser extent. The delicate Crisularia plumosa may also be present. There is a significant echinoderm component in this biotope. Species such as the starfish Asterias rubens, Henricia oculata, Marthasterias glacialis and Luidia ciliaris, the sea urchin Echinus esculentus and the crinoid Antedon bifida are all regularly recorded. Other species which may be observed include the top shell Calliostoma zizyphinium, the colonial ascidian Clavelina lepadiformis and the barnacle Balanus crenatus. The crab Cancer pagurus is typically found under boulders. This variant has been recorded from from various sites including Pembrokeshire, the Calf of Man and the west coast of Ireland.
Depth range
10-20 m, 20-30 m, 30-50 mAdditional information
Please note the molecular taxonomy of the genus Bugula (Fehlauer-Ale et al., 2015) identified several clear genera (clades), Bugula sensu stricto (30 species), Bugulina (24 species), Crisularia (23 species) and the monotypic Virididentulagen. The following review was derived from information concerning species of Bugula prior to their recent revision. The review assumes that, while their taxonomy has changed, the biology of Bugulidae remains similar. Hence, references to Bugula spp. in the text refer to Bugula sensu stricto, Bugulina, and Crisularia species.
Listed By
Sensitivity review
Sensitivity characteristics of the habitat and relevant characteristic species
The CR.HCR.XFa.ByErSp of biotope complex is typically found on wave-exposed circalittoral bedrock or boulders subject to tidal streams ranging from moderately strong to strong, often with a thin layer of silt covering the seabed. Three variants of this biotope have been described; all are characterized by a bryozoan turf with erect sponges. Typical bryozoans include Alcyonidium diaphanum, Flustra foliacea, Pentapora foliacea, Bugulina plumosa and Bugulina flabellata. The biotopes are characterized by a range of erect sponges including Raspailia ramosa, Stelligera stuposa and Stelligera rigida, with others including Cliona celata, Dysidea fragilis, Pachymatisma johnstonia, Polymastia boletiformis, Hemimycale columella, Esperiopsis fucorum (syn. Amphilectus fucorum), Polymastia mamillaris and Tethya aurantium also present Connor et al. (2004). Given limited evidence for the sponges, relevant evidence for non-characterizing sponges is presented where applicable.
ByErSp.DysAct is characterized by the dominance of the sponge Dysidea fragilis and the anemone Actinothoe sphyrodeta and occurs in more tide-swept conditions. ByErSp.Sag is characterized by the additional presence of the anemone Cylista elegans, but occurs in similar conditions to ByErSp.DysAct. There are, however, distinct different geographical distributions. ByErSp.DysAct only tends to be found off Wales and Lundy whereas ByErSp.Cyl is usually found off the west coast of Ireland Connor et al. (2004). ByErSp.Eun is dominated by the seafan Eunicella verrucosa, and is considered separately.
Due to the range of bryozoan and sponge species present, most assessments for this biotope are based on the taxonomic group. Other species present in these biotopes are considered transient, mobile or ubiquitous and are, therefore, not considered significant to the assessment of the sensitivity of this biotope. However, information on the sensitivity of other characterizing species is included where appropriate.
Resilience and recovery rates of habitat
The brooded, lecithotrophic coronate larvae of many bryozoans (e.g. Flustra foliacea, Securiflustra securifrons, and Bugula species), have a short pelagic lifetime of several hours to about 12 hours (Ryland, 1976). Recruitment is dependent on the supply of suitable, stable, hard substrata (Eggleston, 1972b; Ryland, 1976; Dyrynda, 1994). However, even in the presence of available substratum Ryland (1976) noted that significant recruitment in bryozoans only occurred in the proximity of breeding colonies. For example, Hatcher (1998) reported colonization of slabs, suspended 1 m above the sediment, by Bugula fulva within 363 days while Castric-Fey (1974) noted that Bugula turbinata, Bugula plumosa and Bugula calathus did not recruit to settlement plates after ca two years in the subtidal even though present on the surrounding bedrock. Similarly, Keough & Chernoff (1987) noted that Bugula neritina was absent from areas of seagrass bed in Florida even though substantial populations were present <100m away.
Bugula spp. are perennials which tend to form short lived, large colonies in summer with significant die-back in late autumn and a dormant winter phase (Eggleston 1972; Dyrynda & Ryland, 1982). Reproduction occurs in summer/early autumn with some species such as Bugula flabellata reportedly having two generations of fronds capable of reproduction each year (Dyrynda & Ryland, 1982). Eggleston (1972) reported that newly settled specimens from the first generation in the Isle of Man grew rapidly and contributed to the second generation.
Flustra foliacea is a coarse, foliaceous bryozoan that tends to be found on stones and shells, reaches 10 cm in height, is common to all coasts in north-west Europe (Hayward & Ryland, 1995b) and is found across all coasts in the British Isles (NBN, 2016). Stebbing (1974) noted that Flustra foliacea on the Gower peninsular, South Wales had an annual growth season between March and November, with a dormant winter period, when no growth occurred, leading to a line forming across the fronds which can be used to age specimens. The species can regularly reach six years of age, although 12 year old specimens were reported off the Gower Peninsula (Stebbing, 1971; Ryland, 1976). Fortunato et al. (2013) compared numerous sets of growth data with their own observations and reported that colonies grow faster during the first couple of years (about 1.05 cm/year) and slowed down afterwards, which could be due to the lateral growth of the fronds. Colonies appeared to be able to regenerate areas of the frond which had been removed by grazing. Silén (1981) found Flustra foliacea could repair physical damage to its fronds with 5-10 days, concluding that, as long as the holdfast remains intact, Flustra foliacea would survive and grow back. Once settled, new colonies of Flustra foliacea take at least 1 year to develop erect growth and 1-2 years to reach maturity, depending on environmental conditions (Tillin & Tyler Walters, 2014). Four years after sinking off Lundy, the M.V. Roberts was found to be colonized by erect bryozoans and hydroids, including occasional Flustra foliacea (Hiscock, 1981). Flustra foliacea colonies are perennial and potentially highly fecund with increasing colony size as each zooid produces a single embryo (Tillin & Tyler Walters, 2014; Eggleston (1972) with ca 10,000 larvae released from a specimen of Flustra foliacea within 3 hrs (Dalyell, cited in Hincks, 1880). Whilst Bryozoan larvae are typically very short-lived, limiting recruitment to the immediate area surrounding breeding colonies, specimens experiencing strong water movement would improve dispersal potential, and may explain reports of Flustra foliacea colonizing a wreck several hundreds of metres from any significant hard substrata, and hence a considerable distance from potentially parent colonies (Hiscock, 1981; Tyler-Walters & Ballerstedt, 2007). Flustra foliacea requires stable hard substrata (Eggleston, 1972; Ryland, 1976; Dyrynda, 1994) and the abundance of bryozoans is positively correlated with the supply of stable hard substrata and hence with current strength (Eggleston, 1972b; Ryland, 1976).
Alcyonidium diaphanum forms an erect colony that can grow up to 50 cm long but more usually 15 cm. It has a small encrusting base, which attaches to hard substratum. The size, colour and colony form varies widely around the British Isles (Ager, 2007). Pentapora foliacea (syn fascialis)is an erect perennial bryozoan (Eggleston, 1972; Hayward & Ryland, 1995). It recovered in 3.5 years after almost total loss of a local population (Cocito et al., 1998). Pentapora foliacea grows initially as an encrusting sheet, which seems able to regenerate erect growths (P.J. Hayward pers. comm cited in Jackson, 2016). Colonies are typically 20 cm in diameter but can grow up to 2 m in diameter and reach a height of 30 cm in the British Isles (Hayward & Ryland, 1979). Presence or absence of ovicells has been taken to be a reliable indicator of reproductive status and, therefore, of sexual maturity (Cocito et al., 1998). Ovicells were present in colonies in Skomer, Wales in September, indicating a reproduction event in September or late August (Lock et al., 2006). Colonies of Pentapora fascialis as small as 2.8 cm were recorded as having ovicells, with reproduction possible from early stage of colony development (Cocito et al., 1998 cited in Jackson, 2016). Lock et al., 2006 describes growth of Pentapora foliacea in Skomer, Wales as highly variable, with some colonies growing 800 cm² in a year whilst other large colonies completely disappeared. Recovery to pre-disturbance levels following a severe heat event, which resulted in decline of 86% in live colony portion of Pentapora fascialis in the Mediterranean, took four years Cocito & Sgorbini (2014).
Little information on sponge longevity and resilience exists. Reproduction can be asexual (e.g. budding) or sexual (Naylor, 2011) and individual sponges are usually hermaphroditic (Hayward & Ryland, 1994). Short-lived ciliated larvae are released via the aquiferous system and metamorphosis follows settlement. Growth and reproduction are generally seasonal (Hayward & Ryland, 1994). Rejuvenation from fragments can also be considered an important form of reproduction (Fish & Fish, 1996). Marine sponges often harbour dense and diverse microbial communities, which can include bacteria, archaea and single-celled eukaryotes (fungi and microalgae) that comprise up to 40% of sponge volume, and which may have a profound impact on host biology (Webster & Taylor, 2012). Many sponges recruit annually, growth can be quite rapid, with a lifespan of one to several years (Ackers, 1992). However, sponge longevity and growth is highly variable depending on the species and conditions (Lancaster, 2014). It is likely that erect sponges are generally longer lived and slower growing given their more complex nature.
Sponges are known to be highly resilient to physical damage with an ability to survive severe damage, regenerate and reorganize to function fully again (Wulff, 2006). However, the ability to resist damage and recover varies between species (Wulff, 2006). Sponge fragments of coral reef species torn from adults during hurricanes have been observed to re-attach and continue growing (Wulff, 2006). Fowler & Lafoley (1993) monitored marine nature reserves in Lundy and the Isles Scilly and found that a number of more common sponges showed great variation in size and cover during the study period. Large colonies appeared and vanished at some locations. Some large encrusting sponges went through periods of both growth and shrinkage, with considerable changes taking place from year to year. For example, Cliona celata colonies generally grew extremely rapidly, doubling their size or more each year. In some years an apparent shrinkage in size also took place. In contrast, there were no obvious changes in the cover of certain unidentified thin encrusting sponges. Axinellid sponges have been described as very slow growing and little to no recovery has been observed over long periods of monitoring (Fowler & Lafoley, 1993; Hiscock, 1994; 2002). Picton, & Morrow (2015) described Amphilectus fucorum as extremely polymorphic and fast-growing, changing shape in just a few weeks. It may be encrusting as thin sheets or cushions, massive lobose, or branched. Hiscock (pers. comm.) noted that Amphilectus fucorum was found growing on (short-lived) ascidian tests and showed significant seasonal variation in abundance, which suggested this sponge was highly resilient. Dysidea fragilis readily colonizes deep water wrecks (Ackers et al., 1992) Tethya aurantium produces stalked reproductive buds between July and September (van Soest et al., 2000). Raspailia ramosa, a branching sponge, spawns in September (Lévi, 1956, cited from Van Soest, 2000). Stelligera stuposa is a branching erect sponge commonly found in Britain’s circalittoral (Picton & Morrow, 2015). Polymastia mamilliaris is an encrusting sponge present from the Arctic to the Mediterranean (Boury-Esnault, 1987). Polymastia boletiformis is a commonly found spherical sponge found across the Western and Eastern Atlantic and is present from the Arctic to the Mediterranean (Boury-Esnault, 1987). Pachymatisma johnstonia can be massive-lobose, hemispherical to irregularly rounded and up to 30 cm or more across (Picton & Morrow, 2015). Sebens (1985, 1986) found that the sponge Halichondria panicea reached pre-clearance levels of cover after 2 years. A few individuals of Alcyonium digitatum and Metridium senile colonized within 4 years (Sebens, 1986) and slower-growing sponges would probably take longer to reach pre-clearance levels.
Sargatia elegans is found from Scandinavia to the Mediterranean (Picton & Morrow 2016). Actinothoe sphyrodeta is distributed from the northern coast of Scotland to Spain (Ramos, 2010; NBN, 2016). However, no information on resilience was found for these species. Actinothoe sphyrodeta was described as rapidly maturing (within 1 year) with a low fecundity (Jackson et al., 2008).
Anemones are not sessile but are capable of slow movement. For example, Sebens (1981) observed immigration to cleared patches of the long-lived anemone Anthopleura xanthogrammica as being the primary driver towards recovery. Sebens (1981) cleared intertidal patches of Anthopleura xanthogrammica at Mukkaw Bay, WA observing the effects over four years. Even after four years cleared areas were not back to pre-removal population densities. Chia & Spaulding (1972) studying the anemone Tealia crassicornis found no sign of gonad development at 14 months old. Wedi & Dunn (1983) recorded the smallest fertile Urticina lofotensis at 18 months old.
The recolonization of epifauna on vertical rock walls was investigated by Sebens (1985, 1986). He reported that rapid colonizers such as encrusting corallines, encrusting bryozoans, amphipods and tubeworms recolonized within 1-4 months. Ascidians such as Dendrodoa carnea, Molgula manhattensis and Aplidium spp. achieved significant cover in less than a year, and, together with Halichondria panicea, reached pre-clearance levels of cover after 2 years. A few individuals of Alcyonium digitatum and Metridium senile colonized within 4 years (Sebens, 1986) and would probably take longer to reach pre-clearance levels.
Jensen et al. (1994) reported the colonization of an artificial reef in Poole Bay, England. They noted that erect bryozoans, including Bugula plumosa, began to appear within 6 months, reaching a peak in the following summer, 12 months after the reef was constructed. Similarly, ascidians colonized within a few months e.g. Aplidium spp. Sponges were slow to establish with only a few species present within 6-12 months but beginning to increase in number after 2 years, while anemones were very slow to colonize with only isolated specimens present after 2 years (Jensen et al., 1994.). In addition, Hatcher (1998) reported a diverse mobile epifauna after a year’s deployment of her settlement panels.
Long-term studies of fixed quadrats in epifaunal communities demonstrated that while seasonal and annual changes occurred, subtidal faunal turf communities were relatively stable, becoming more stable with increasing depth and substratum stability (i.e. bedrock and large boulders rather than small rocks) (Osman, 1977; Hartnoll, 1998). Many of the faunal turf species are long-lived, e.g. 6 -12 years in Flustra foliacea, 5-8 years in Ascidia mentula, over 20 years in Alcyonium digitatum, 8-16 years in Echinus esculentus and probably many hydroids (Stebbing, 1971a; Gili & Hughes, 1995; Hartnoll, 1998). However, Bugula dominated communities recorded of west Anglesey in 1996 were reported to be 'silted and ragged' in the same season the following year, suggesting some inter-annual variation may occur (Brazier et al., 1999).
Resilience assessment. The majority of the species within this biotope have short-lived pelagic larvae, with limited powers of dispersal, resulting in good local recruitment but poor long-distance dispersal. Bugula and other bryozoan and hydroid species exhibit multiple generations per year, that involve good local recruitment, rapid growth and reproduction. Bryozoans and hydroids are often opportunistic, fouling species, that colonize and occupy space rapidly. For example, hydroids would probably colonize within 1-3 months and return to their original cover rapidly, while Bugula species have been reported to colonize new habitats within 6 -12 months (see recruitment). However, Bugula has been noted to be absent from available habitats even when large populations are nearby (Castric-Frey, 1974; Keough & Chernoff, 1987), suggesting that recruitment may be more sporadic. Where the population is reduced in extent or abundance but individuals remain, local recruitment, augmented by dormant resistant stages and asexual reproduction, is likely to result in rapid recovery of the dominant bryozoan species, hydroids, probably within 12 months. Colonial ascidians would probably recover their original cover within two years, while sponges and anemones may take longer to recover but would probably regain their original cover within five years. Where the community was destroyed and recovery is dependent on recruitment from other areas, bryozoans, hydroids and ascidians would probably recruit rapidly from other neighbouring areas (see Jensen et al., 1994; Hatcher, 1998). However, sponges and especially Anthozoa may take many years to recruit and develop. Sebens (1985, 1986) found that the sponge Halichondria panicea reached pre-clearance levels of cover after 2 years. A few individuals of Alcyonium digitatum and Metridium senile colonized within 4 years (Sebens, 1986) and slower-growing sponges would probably take longer to reach pre-clearance levels. Some of the faster-growing, smaller sponges would colonize new sites relatively quickly, little information regarding the resilience of larger, branching sponges was sparse, and a more cautious resilience assessment is therefore applied.
Overall, if the community suffers significant mortality from a pressure (resistance of ‘None’, ‘Low’) resilience is assessed as ‘Medium’ (recovery within 2-10 years). If resistance is assessed as ‘Medium’ or ‘High’ then resilience will be assessed as ‘High’ (recovery within 2 years). Confidence is assessed as ‘Low’, given the lack of resilience, growth rates or fecundity for characterizing sponges. The anemones are likely to be longer lived and take longer to recover.
Hydrological Pressures
Use [show more] / [show less] to open/close text displayed
| Resistance | Resilience | Sensitivity | |
Temperature increase (local) [Show more]Temperature increase (local)Benchmark. A 5°C increase in temperature for one month, or 2°C for one year. Further detail EvidenceBugula spp.grow and reproduce in the summer months, however, day length and/or the phytoplankton bloom characteristic of temperate waters are probably more important than temperature (Ryland, 1967; 1970; Tyler-Walters, 2005). Bugula turbinatais a predominantly southern species in British waters (Lewis, 1964; Hayward & Ryland, 1998) but has been recorded as far north as Shetland (NBN, 2016). A long-term increase in temperature may increase its abundance in northern British waters and allow the species to extend its range. It occurs as far south as the Mediterranean (Rosso, 2003) and is likely to tolerate increases of temperature, at the benchmark level. Cocito & Sgorbini (2014) studied spatial and temporal patterns of colonial bryozoans in the Ligurian Sea over 9 years. High temperature events were recorded. The first caused mass mortality among a number of species. Decline in Pentapora fascialis colony cover between 11 and 22 m depth followed an unusually warm summer in 1999 (with larger colonies most affected. The temperature at 11 m was 23.87 ± 1.4 °C resulted in mortality of 86% of a colony. Recovery took place gradually, with deeper communities recovering to pre-disturbance levels within four years. Pentapora foliacea is found in warmer waters as far south the north coast of Morocco, with the northernmost limits of its distribution in the Minch off western Scotland (Lombardi et al., 2010). Once established, colonies are most likely able to withstand occasional lower or higher than normal temperatures, but long-term decreases in temperature may cause distribution range to shrink. Although being quite long-lived (10+ years) (Jackson, 2016), Alcyonidium diaphanum is commonly found across the British Isles and is probably widely distributed across North-West Europe (Fish & Fish, 1992). The sponges, Tethya aurantium, Dysidea fragalis are found from the Arctic to the Mediterranean, Amphilectus fucorum from Norway to France, Polymastia boletiformis from the Arctic to the Atlantic coasts of Europe, and Raspailia ramosa is found across the western British Isles and Northern France (Ackers, 1993). The sea anemone Sargatia elegans is found from Scandinavia to the Mediterranean (Picton & Morrow 2016) and Actinothoe sphyrodeta is distributed from the northern coast of Scotland to Spain (Ramos, 2010; NBN, 2016). Berman et al. (2013) monitored sponge communities off Skomer Island, UK over three years with all characterizing sponges for this biotope assessed. seawater temperature, turbidity, photosynthetically active radiation and wind speed were all recorded during the study. It was concluded that, despite changes in species composition, primarily driven by the non-characterizing Hymeraphia, Stellifera and Halicnemia patera, no significant difference in sponge density was recorded in all sites studied. Morphological changes most strongly correlated with a mixture of water visibility and temperature. Cebrian et al. (2011) conducted four-year surveys of two shallow-water sponge species, Ircinia fasciculata and Sarcotragus spinosulum in the western Mediterranean Sea. Two severe sponge die-offs (total mortality ranging from 80 to 95% of specimens) occurred in the summers of 2008 and 2009. These events primarily affected Ircinia fasciculata, and a significant positive correlation was observed between elevated temperature and injured specimens. It was suggested, following in vitro studies of the associated cyanobacteria in increasing temperatures up to those experienced in ‘extreme summer’ of 27°C, that heat related disappearance of the cyanobacteria in Ircinia fasciculata (a bacteriosponge) was important when considering sponge mortality. Research by Webster et al.(2008, 2011), Webster & Taylor, (2012), Preston &Burton (2015) suggested that many sponges rely on a holobiont of many synergistic microbes. Webster et al., 2011 describes a much higher thermal tolerance to sponge larval holobiont when compared with adult sponges. Adult Rhopaloeides odorabile from the Great Barrier Reef has been shown to have a strict thermal threshold of between 31-33°C (Webster et al. 2008) whereas the larvae could tolerate temperatures of up to 36°C with no adverse effects. Sensitivity assessment. The characterizing species are widely distributed across the British Isles, none being at their southern limit. Morphological changes were observed in UK sponge communities, with temperature a factor, but the characterizing sponges assessed were not listed as the most highly contributing to these changes (Berman et al., 2013). Resistance has been assessed as ‘High’, resilience as ‘High’ and sensitivity as ‘Not Sensitive’ at the benchmark level. | HighHelp | HighHelp | Not sensitiveHelp |
Temperature decrease (local) [Show more]Temperature decrease (local)Benchmark. A 5°C decrease in temperature for one month, or 2°C for one year. Further detail EvidenceAll characterizing bryozoans (Alcyonidium diaphanum, Flustra foliacea, Pentapora foliacea, Bugula plumosa and Bugula flabellata) have been recorded across the British Isles, from the Channel Isles to the northern coast of Scotland (NBN, 2015). Patzold et al. (1987) recorded the formation of a growth band in Pentapora foliacea during times of reduced reproduction, which appeared during periods of colder water temperatures. Pentapora foliacea is found in warmer waters as far south as the northern coast of Morocco, with the northernmost limits of distribution in the Minch off western Scotland (Lombardi et al., 2010). The sponges Tethya aurantium and Dysidea fragalis are found from the Arctic to the Mediterranean, Amphilectus fucorum is found from Norway to France, Polymastia boletiformis is found from the Arctic to the Atlantic coasts of Europe, Raspailia ramosa found across the western British Isles and Northern France (Ackers, 1993). Berman et al. (2013) monitored sponge communities off Skomer Island, UK over three years with all characterizing sponges for this biotope assessed. seawater temperature, turbidity, photosynthetically active radiation and wind speed were all recorded during the study. It was concluded that, despite changes in species composition, primarily driven by the non-characterizing Hymeraphia Stellifera and Halicnemia patera, no significant difference in sponge density was recorded in all sites studied. Morphological changes most strongly correlated with a mixture of visibility and temperature. Some sponges do exhibit morphological strategies to cope with winter temperatures e.g. Halichondria bowerbanki goes into a dormant state below 4°C, characterized by major disintegration and loss of choanocyte chambers with many sponges surviving mild winters in more protected areas from where it can recolonize (Vethaak et al., 1982). Crisp (1964) reported the effects of an unusually cold winter (1962-3) on the marine life in Britain, including Porifera in North Wales. Whilst difficulty distinguishing between mortality and delayed development was noted, Crisp (1964) found that Pachymastia johnstonia and Halichondria panicea were wholly or partly killed by frost, several species appeared to be missing including Amphilectus fucorum. Others, including Hymeniacidon perleve were unusually rare and a few species, including Polymstia boletiformis, were not seriously affected. It should be noted that Crisp’s general comments on all marine life state that damage decreased the deeper the habitat. The anemone Sargatia elegans is found from Scandinavia to the Mediterranean (Picton & Morrow 2016), while Actinothoe sphyrodeta is distributed from the northern coast of Scotland to Spain (Ramos, 2010; NBN, 2016) and could therefore be affected by a reduction in temperature. Sensitivity assessment There is evidence of sponge mortality at extreme low temperatures in the British Isles. Given this evidence, it is likely that a cooling of 5°C would affect some of the characterizing sponges, and resistance is assessed as ‘Medium’. A resilience of ‘High’ is recorded and sensitivity is assessed as ‘Low’. | MediumHelp | HighHelp | LowHelp |
Salinity increase (local) [Show more]Salinity increase (local)Benchmark. A increase in one MNCR salinity category above the usual range of the biotope or habitat. Further detail EvidenceMarin (1997) describes the presence of Dysidea fragilis in a hypersaline coastal lagoon (42-47 g/l) in La Mar Menor, Spain. No other evidence could be found for characterizing sponges. Soule & Soule (1979) cite Hastings (1927) who described the presence of five bryozoans in hypersaline conditions in the Suez Canal. Both Cylista elegans and Actinothoe sphyrodeta occur in the littoral (Picton & Morrow, 2016) and are therefore likely to experience both higher and lower salinities than ‘Full’ (30-35 ppt) as per the biotope description (Connor et al., 2004). Sensitivity assessment. The ByErSp biotope complex is a circalittoral biotope that experiences full salinity. Whilst there is some evidence of some bryozoan and sponge species existing in hypersaline conditions, ‘No evidence' could be found for the characterizing bryozoans or the majority of sponge species. | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Salinity decrease (local) [Show more]Salinity decrease (local)Benchmark. A decrease in one MNCR salinity category above the usual range of the biotope or habitat. Further detail EvidenceRyland (1970) stated that, with a few exceptions, the Gymnolaemata were fairly stenohaline and restricted to full salinity (30-35 ppt), noting that reduced salinities result in an impoverished bryozoan fauna. Flustra foliacea appears to be restricted to areas with high salinity (Tyler-Walters & Ballerstedt, 2007; Budd 2008). Dyrynda (1994) noted that Flustra foliacea and Alcyonidium diaphanum were probably restricted to the vicinity of the Poole Harbour entrance by their intolerance to reduced salinity. Although, protected from extreme changes in salinity due to their subtidal habitat, severe hyposaline conditions could adversely affect Flustra foliacea colonies. Castric & Chassé (1991) conducted a factorial analysis of the subtidal rocky ecology near Brest, France and rated the distribution of species from estuarine to offshore conditions. Dysidea fragilis and Raspailia ramosa were rated as indifferent to this range. Cliona celata and Pachymatisma johnstonia had a slight preference for more estuarine conditions while Polymastia mamillaris and Tethya aurantium had a slight preference for offshore conditions. Stelligera rigida and Polymastia boletiformis (as Polymastia robusta) were intolerant of the more estuarine conditions. Mean salinity difference between the two farthest zones was low (35.1 and 33.8 ‰ respectively) but with a greater range being experienced in the Inner Rade (± 0.1 compared with 2.4‰). It should be noted that the range of salinities identified in this study do not reach the lower benchmark level, and at least some of the characterizing sponges are likely to be affected at the benchmark level. Some characterizing sponges are present in lower salinity biotopes, such as CR.MCR.CFaVS (Connor et al., 2004) and proportion of the sponge community is likely to survive a low salinity event. Both Cylista elegans and Actinothoe sphyrodeta occur in the littoral (Picton & Morrow, 2016) and are therefore likely to experience both higher and lower salinities than ‘Full’ (30-35 ppt) as per the biotope description (Connor et al., 2004). Sensitivity assessment. Some of the characterizing bryozoans and sponges are likely to be adversely affected by a reduction in salinity. Resistance is assessed as ‘Low’, resilience as ‘Medium’ and sensitivity as ‘Medium’. | LowHelp | MediumHelp | MediumHelp |
Water flow (tidal current) changes (local) [Show more]Water flow (tidal current) changes (local)Benchmark. A change in peak mean spring bed flow velocity of between 0.1 m/s to 0.2 m/s for more than one year. Further detail EvidenceWater flow has been shown to be important for the development of bryozoan communities and the provision of suitable hard substrata for colonization (Eggleston, 1972b; Ryland, 1976). In addition, areas subject to high mass transport of water such as the Menai Strait and tidal rapids generally support large numbers of bryozoan species (Moore, 1977). Although, active suspension feeders, their feeding currents are probably fairly localized and they are dependent on water flow to bring adequate food supplies within reach (McKinney, 1986). A substantial decrease in water flow will probably result in impaired growth due to a reduction in food availability, and an increased risk of siltation (Tyler-Walters, 2005). Okamura (1984) reported that an increase in water flow from slow flow (1-2 cm/s) to fast flow (10-12 cm/s) reduced feeding efficiency in small colonies but not in large colonies of Bugula stolonifera. Flustra foliacea colonies are flexible, robust and reach high abundances in areas subject to strong currents and tidal streams Stebbing, 1971; Eggleston, 1972b; Knight-Jones & Nelson-Smith, 1977; Hiscock, 1983, 1985; Holme & Wilson, 1985). Dyrynda (1994) suggested that mature fronded colonies do not occur on unstable substratum due to the drag caused by their fronds, resulting in rafting of colonies on shells or the rolling of pebbles and cobbles, and destruction of the colony. Dyrynda (1994) reported that the distribution of Flustra foliacea in the current swept entrance to Poole Harbour was restricted to circalittoral boulders, on which it dominated as nearly mono-specific stands. While, the pumping activity of the lophophores provide the greatest proportion of the colonies food requirements (Hayward & Ryland, 1998), the current generated is probably very localized and the colonies are likely to be dependent on water currents for food supply. A significant decrease in water flow is likely to result in a decrease in the abundance of bryozoans. Riisgard et al., 1993 discussed the low energy cost of filtration for sponges and concluded that passive current-induced filtration may be of insignificant importance for sponges. Pumping and filtering occurs in choanocyte cells which generate water currents in sponges using flagella (de Vos et al., 1991). Sensitivity assessment The biotope complex CR.HCR.XFa.ByErSp is defined as a moderate to high energy and occurs in areas of moderately strong to strong tidal water flow (1-6 kn). Bryozoan communities rely on movement of water for feeding and a severe reduction in water flow over an extended period of time could cause mortality. Sponges are present in biotopes with both stronger and negligible tidal flow and are therefore unlikely to be affected by a change in water flow. However, at the benchmark level (0.1-0.2 m/s), it is unlikely that an increase or decrease would increase chances of mortality for any of the characterizing species or require a reclassification of the biotope. Resistance is therefore recorded as ‘High’ with resilience as ‘High’ and the biotope is ‘Not sensitive’ at the benchmark level. | HighHelp | HighHelp | Not sensitiveHelp |
Emergence regime changes [Show more]Emergence regime changesBenchmark. 1) A change in the time covered or not covered by the sea for a period of ≥1 year or 2) an increase in relative sea level or decrease in high water level for ≥1 year. Further detail EvidenceChanges in emergence are not relevant to this biotope as it is restricted to fully subtidal/circalittoral conditions - the pressure benchmark is relevant only to littoral and shallow sublittoral fringe biotopes. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Wave exposure changes (local) [Show more]Wave exposure changes (local)Benchmark. A change in near shore significant wave height of >3% but <5% for more than one year. Further detail EvidenceBugula spp. produce flexible erect tufts, which are likely to move with the oscillatory flow created by wave action. Bugula turbinata has been recorded from very wave exposed to very wave sheltered habitats (Tyler-Walters, 2005). Flustra foliacea occurs from very wave exposed to sheltered waters, although probably limited to deeper waters in very wave exposed conditions (Tyler-Walters & Ballerstedt ,2007) The oscillatory water flow generated by wave action may be more damaging than constant strong currents, e.g. strong wave action may generate an oscillatory flow of 2 m/sec at 20 m (Hiscock, 1983, 1985). Flustra foliacea is a common member of the flotsam, having been removed from its substratum by storms.. Whilst the biotope is circalittoral, a severe increase in wave exposure (e.g. storms) could affect bryozoans colonies. Cocito et al (1998) described a severe winter storm of 1993 had devastating effects on a Flustra foliacea population, sweeping away most of the colonies down to 11 m. Roberts et al. (2006) studied deep sponge reef communities (18-20 m) in sheltered and exposed locations in Australia. They reported greater diversity and cover (>40% cover) of sponges in wave-sheltered areas compared with a sparser and more temporal cover in exposed sites (25% cover). Sensitivity assessment The ByEuSp complex is found in extreme wave exposure so that a further increase is Not relevant. However, a reduction in wave exposure is likely to result in faunal communities typical of moderate to low energy, and less wave exposed, habitats, e.g. BrAs complex dominated by ascidians and brittlestars, Flustra, or echinoderm grazed faunal turfs. Hence, a significant reduction in wave exposure could result in reclassification and loss of the biotope. However, a 3-5% change in significant wave height (the benchmark) is probably not significant. Resistance is, therefore, recorded as ‘High’ with resilience as ‘High’ and the biotope is ‘Not sensitive’ at the benchmark level. | HighHelp | HighHelp | Not sensitiveHelp |
Chemical Pressures
Use [show more] / [show less] to open/close text displayed
| Resistance | Resilience | Sensitivity | |
Transition elements & organo-metal contamination [Show more]Transition elements & organo-metal contaminationBenchmark. Exposure of marine species or habitat to one or more relevant contaminants via uncontrolled releases or incidental spills. Further detail EvidenceBryozoans are common members of the fouling community, and amongst those organisms most resistant to antifouling measures, such as copper containing anti-fouling paints (Soule & Soule, 1979; Holt et al., 1995). Bryan & Gibbs (1991) reported that there was little evidence regarding TBT toxicity in bryozoa with the exception of the encrusting Schizoporella errata, which suffered 50% mortality when exposed for 63 days to 100ng/l TBT. Rees et al. (2001) reported that the abundance of epifauna (including bryozoans) had increased in the Crouch estuary in the 5 years since TBT was banned from use on small vessels. This last report suggests that bryozoans may be at least inhibited by the presence of TBT. This pressure is Not assessed but evidence is presented where available. | Not Assessed (NA)Help | Not assessed (NA)Help | Not assessed (NA)Help |
Hydrocarbon & PAH contamination [Show more]Hydrocarbon & PAH contaminationBenchmark. Exposure of marine species or habitat to one or more relevant contaminants via uncontrolled releases or incidental spills. Further detail EvidenceThis pressure is Not assessed but evidence is presented where available. ByErSp is a sub-tidal biotopecomplex (Connor et al., 2004). Oil pollution is mainly a surface phenomenon its impact upon circalittoral turf communities is likely to be limited. However, as in the case of the Prestige oil spill off the coast of France, high swell and winds can cause oil pollutants to mix with the seawater and potentially negatively affect sub-littoral habitats (Castège et al., 2014). Filter feeders are highly sensitive to oil pollution, particularly those inhabiting the tidal zones which experience high exposure and show correspondingly high mortality, as are bottom dwelling organisms in areas where oil components are deposited by sedimentation (Zahn et al., 1981). There is little information on the effects of hydrocarbons on bryozoans. Ryland & Putron (1998) did not detect adverse effects of oil contamination on the bryozoan Alcyonidium spp. in Milford Haven or St. Catherine's Island, south Pembrokeshire, although it did alter the breeding period. Banks & Brown (2002) found that exposure to crude oil significantly impacted recruitment in the bryozoan Membranipora savartii. Tethya lyncurium concentrated BaP (benzo[a ]pyrene )to 40 times the external concentration and no significant repair of DNA was observed in the sponges, which, in higher animal, would likely lead to cancers. As sponge cells are not organized into organs the long-term effects are uncertain (Zahn et al., 1981). | Not Assessed (NA)Help | Not assessed (NA)Help | Not assessed (NA)Help |
Synthetic compound contamination [Show more]Synthetic compound contaminationBenchmark. Exposure of marine species or habitat to one or more relevant contaminants via uncontrolled releases or incidental spills. Further detail EvidenceHoare & Hiscock (1974) suggested that polyzoa (bryozoa) were amongst the most intolerant species to acidified halogenated effluents in Amlwch Bay, Anglesey and reported that Flustra foliacea did not occur less than 165m from the effluent source. The evidence therefore suggests that Securiflustra securifrons would be sensitive to synthetic compounds. | Not Assessed (NA)Help | Not assessed (NA)Help | Not assessed (NA)Help |
Radionuclide contamination [Show more]Radionuclide contaminationBenchmark. An increase in 10µGy/h above background levels. Further detail Evidence'No evidence'. | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Introduction of other substances [Show more]Introduction of other substancesBenchmark. Exposure of marine species or habitat to one or more relevant contaminants via uncontrolled releases or incidental spills. Further detail EvidenceThis pressure is Not assessed. | Not Assessed (NA)Help | Not assessed (NA)Help | Not assessed (NA)Help |
De-oxygenation [Show more]De-oxygenationBenchmark. Exposure to dissolved oxygen concentration of less than or equal to 2 mg/l for one week (a change from WFD poor status to bad status). Further detail EvidenceIn general, respiration in most marine invertebrates does not appear to be significantly affected until extremely low concentrations are reached. For many benthic invertebrates this concentration is about 2 ml/l (Herreid, 1980; Rosenberg et al., 1991; Diaz & Rosenberg, 1995). Cole et al. (1999) suggest possible adverse effects on marine species below 4 mg/l and probable adverse effects below 2 mg/l. Little information on the effects of oxygenation on bryozoans was found. Sagasti et al. (2000) reported that epifauna communities, including dominant species such as the bryozoans were unaffected by periods of moderate hypoxia (ca 0.35 -1.4 ml/l which corresponds to ca 0.5 - 2 mg/l ) and short periods of hypoxia (<0.35 ml/l which corresponds to <0.5 mg/l) in the York River, Chesapeake Bay, although bryozoans were more abundant in the area with generally higher oxygen. However, estuarine species are likely to be better adapted to periodic changes in oxygenation. Hiscock & Hoare (1975) reported an oxycline forming in the summer months (Jun-Sep) in a quarry lake (Abereiddy, Pembrokeshire) from close to full oxygen saturation at the surface to <5% saturation (ca <0.5 mg/l) below ca 10 m. No Tethya aurantia, Kirchenpaueria pinnata, Hymeniacidon pereleve or Polymastia boletiformis were recorded at depths below 10 - 11 m. Demosponges maintained under laboratory conditions can tolerate hypoxic conditions for brief periods, (Gunda & Janapala, 2009) investigated the effects of variable dissolved oxygen (DO) levels on the survival of the marine sponge, Haliclona pigmentifera. Under hypoxic conditions (1.5-2.0 ppm DO which corresponds to ca 1.5-2.0 mg/l), Haliclona pigmentifera with intact ectodermal layers and subtle oscula survived for 42 ± 3 days. Sponges with prominent oscula, foreign material, and damaged pinacoderm exhibited poor survival (of 1-9 days) under similar conditions. Complete mortality of the sponges occurred within 2 days under anoxic conditions (<0.3 ppm DO = ca <0.3 mg/l). Sensitivity assessment: The evidence suggests that the sponge communities would be severely affected in hypoxic conditions. Resistance is therefore recorded as ‘Low’, with a resilience of ‘Medium’ and sensitivity is classed as 'Medium' at the benchmark level. | LowHelp | MediumHelp | MediumHelp |
Nutrient enrichment [Show more]Nutrient enrichmentBenchmark. Compliance with WFD criteria for good status. Further detail EvidenceHartikainen et al. (2009) reported that increased nutrient concentrations resulted in freshwater bryozoans achieving higher biomass. O’Dea & Okamura (2000) found that annual growth of Flustra foliacea in western Europe substantially increased since 1970. They suggested that this could be due to eutrophication in coastal regions due to organic pollution, leading to increased phytoplankton biomass (see Allen et al., 1998). Gochfeld et al. (2012) studied the effect of nutrient enrichment (≤0.05 to 0.07 μM for nitrate and ≤0.5 μM for phosphate) as a potential stressor in Aplysina caulifornis and its bacterial symbionts, and found that nutrient enrichment had no effects on sponge or symbiont physiology when compared to control . This study does contradict findings in Gochfeld et al. (2007) in which Aplysina spp. sponges were virtually absent from a site of anthropogenic stress in Bocas del Toro, Panama which experienced high rainfall and terrestrial runoff. The author suggested that whilst this site did include elevated nutrient concentrations, other pressures and stresses could be contributing. Rose & Risk, 1985 described increase in abundance of Cliona delitrix in organically polluted section of Grand fringing reef affected by the discharge of untreated faecal sewage. Ward-Paige et al., 2005 described the greatest size and biomass of clionids corresponded with highest nitrogen, ammonia and ɗ15N levels. This biotope is considered to be 'Not sensitive' at the pressure benchmark, that assumes compliance with good status as defined by the WFD. | Not relevant (NR)Help | Not relevant (NR)Help | Not sensitiveHelp |
Organic enrichment [Show more]Organic enrichmentBenchmark. A deposit of 100 gC/m2/yr. Further detail EvidenceO’Dea & Okamura (2000) found that annual growth of Flustra foliacea in western Europe has substantially increased since 1970. They suggest that this could be due to eutrophication in coastal regions due to organic pollution, leading to increased phytoplankton biomass (see Allen et al., 1998). Rose & Risk, 1985 described increase in abundance of the sponge Cliona delitrix in an organically polluted section of Grand Cayman fringing reef affected by the discharge of untreated faecal sewage. De Goeij et al. (2008) used 13C to trace the fate of dissolved organic matter in the coral reef sponge Halisarca caerulea. Biomarkers revealed that the sponge incorporated dissolved organic matter through both bacteria mediated and direct pathways, suggesting that it feeds, directly and indirectly, on dissolved organic matter. Mayer-Pinto & Junqueira (2003) studied the effects of organic pollution on fouling communities in Brazil and found that tolerance of polluted/unpolluted artificial reefs varied among bryozoan species. It should be noted that Bugula spp. preferred the polluted sites. Sensitivity assessment The evidence suggests that organic enrichment may allow some species in the biotope to increase in abundance. Therefore, resistance to this pressure is assessed as 'High' and resilience as 'High'. This biotope is therefore considered to be 'Not sensitive'.
| HighHelp | HighHelp | Not sensitiveHelp |
Physical Pressures
Use [show more] / [show less] to open/close text displayed
| Resistance | Resilience | Sensitivity | |
Physical loss (to land or freshwater habitat) [Show more]Physical loss (to land or freshwater habitat)Benchmark. A permanent loss of existing saline habitat within the site. Further detail EvidenceAll marine habitats and benthic species are considered to have a resistance of ‘None’ to this pressure and to be unable to recover from a permanent loss of habitat (resilience is ‘Very low’). Sensitivity within the direct spatial footprint of this pressure is therefore ‘High’. Although no specific evidence is described confidence in this assessment is ‘High’, due to the incontrovertible nature of this pressure. | NoneHelp | Very LowHelp | HighHelp |
Physical change (to another seabed type) [Show more]Physical change (to another seabed type)Benchmark. Permanent change from sedimentary or soft rock substrata to hard rock or artificial substrata or vice-versa. Further detail EvidenceIf rock were replaced with sediment, this would represent a fundamental change to the physical character of the biotope and the species would be unlikely to recover. The biotope would be lost. Sensitivity assessment. Resistance to the pressure is considered ‘None’, and resilience ‘Very low’. Sensitivity has been assessed as ‘High’. | NoneHelp | Very LowHelp | HighHelp |
Physical change (to another sediment type) [Show more]Physical change (to another sediment type)Benchmark. Permanent change in one Folk class (based on UK SeaMap simplified classification). Further detail Evidence‘Not relevant’ to biotopes occurring on bedrock. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Habitat structure changes - removal of substratum (extraction) [Show more]Habitat structure changes - removal of substratum (extraction)Benchmark. The extraction of substratum to 30 cm (where substratum includes sediments and soft rock but excludes hard bedrock). Further detail EvidenceThe species characterizing this biotope are epifauna or epiflora occurring on rock and would be sensitive to the removal of the habitat. However, extraction of rock substratum is considered unlikely and this pressure is considered to be ‘Not relevant’ to hard substratum habitats. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Abrasion / disturbance of the surface of the substratum or seabed [Show more]Abrasion / disturbance of the surface of the substratum or seabedBenchmark. Damage to surface features (e.g. species and physical structures within the habitat). Further detail EvidencePhysical disturbance by fishing gear has been shown to adversely affect emergent epifaunal communities with hydroid and bryozoan matrices reported to be greatly reduced in fished areas (Jennings & Kaiser, 1998). Heavy mobile gears could also result in movement of boulders (Bullimore, 1985; Jennings & Kaiser, 1998). Picton & Morrow (2015) describe Amphilectus fucorum as extremely polymorphic and fast growing, changing shape in just a few weeks. It may be encrusting as thin sheets or cushions, massive lobose, with or without tassels, or branched. It has a soft texture and open surface and is therefore likely to be damaged through abrasion pressures. Picton & Morrow (2015) noted that whilst Raspailia ramosa is firm and elastic, the soft layer is easily rubbed from the strong axial core. Freese et al. (1999) studied the effects of trawling on seafloor habitats and associated invertebrates in the Gulf of Alaska. They found that a transect following a single trawling event showed significantly reduced ‘vase’ sponges (67% expressed damage), ‘morel’ sponges (total damage could not be quantified as their brittle nature meant that these sponges were completely torn apart and scattered). The ‘finger’ sponges, the smallest and least damaged of the sponges assessed, were damaged by being knocked over (14%). Van Dolah et al. (1987) studied the effects on sponges and corals of one trawl event over a low-relief hard bottom habitat off Georgia, US. The densities of individuals taller than 10 cm of three species of sponges in the trawl path and in adjacent control area were assessed by divers, and were compared before, immediately after and 12 months after trawling. Of the total number of sponges remaining in in the trawled area, 32% were damaged. Most of the affected sponges were the barrel sponges Cliona spp., whereas Haliclona oculta and Ircina campana were not significantly affected. Twelve months after trawling, the abundance of sponges had increased to pre-trawl densities, or greater. Tilnant (1979) found that, following a shrimp trawl in Florida, US, over 50% of sponges, including Neopetrosia, Spheciospongia, Spongia and Hippiospongia, were torn loose from the bottom. Highest damage incidence occurred to the finger sponge Neopetrosia longleyi. Size did not appear to be important in determining whether a sponge was affected by the trawl. Recovery was ongoing, but not complete 11 months after the trawl, although no specific data was provided. Freese (2001) studied deep cold-water sponges in Alaska a year after a trawl event; 46.8% of sponges exhibited damage with 32.1% having been torn loose. None of the damaged sponges displayed signs of regrowth or recovery. This was in stark contrast to early work by Freese (1999) on warm shallow sponge communities. Impacts of trawling activity in Alaska study were more persistent due to the slower growth/regeneration rates of deep, cold-water sponges. Given the slow growth rates and long lifespans of the rich, diverse fauna, it was considered likely to take many years for deep sponge communities to recover if adversely affected by physical damage Freese (2001). Boulcott & Howell (2011) conducted experimental Newhaven scallop dredging over a circalittoral rock habitat in the sound of Jura, Scotland and recorded the damage to the resident community. The results indicated that vulnerable epifauna, including the sponge Pachymatisma Johnstoni, were highly damaged by the experimental trawl. It should be noted that other epifaunal turfs on uneven rock substrata were more resistant to damage than populations on sediment. Coleman et al., 2013 described a 4 year study on the differences between a commercially potted area in Lundy with a no take zone. No significant difference in Axinellid populations was observed. The authors concluded that lighter abrasion pressures, such as potting, were far less damaging than heavier gears, such as trawls. Whilst capable of movement, the anemones are not rapid and are likely to be as exposed to abrasion events as the sessile epifauna. Anemones are also soft bodied and are likely to be intolerant of abrasion events. Sensitivity assessment Given the sessile, emerged nature of the sponges and bryozoans, damage and mortality following a physical disturbance effect are likely to be significant, however some studies have brought into question the extent of damage to the faunal turf. The physiology of the bryozoans affords some protection in the event of abrasion events and recovery is likely to be rapid if stolons remain undamaged. However, b ased on the damage to sponges, resistance has been assessed as ‘Low’, resilience as ‘Medium’ and sensitivity has been assessed as ‘Medium’. Please note Boulcott & Howell (2011) did not mention the abrasion caused by fully loaded collection bags on the Newhaven dredges. A fully loaded Newhaven dredge may cause higher damage to community as indicated in their study. | LowHelp | MediumHelp | MediumHelp |
Penetration or disturbance of the substratum subsurface [Show more]Penetration or disturbance of the substratum subsurfaceBenchmark. Damage to sub-surface features (e.g. species and physical structures within the habitat). Further detail EvidenceThe species characterizing this biotope group are epifauna or epiflora occurring on rock which is resistant to subsurface penetration. The assessment for abrasion at the surface only is therefore considered to equally represent sensitivity to this pressure. This pressure is thought ‘Not Relevant’ to hard rock biotopes | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Changes in suspended solids (water clarity) [Show more]Changes in suspended solids (water clarity)Benchmark. A change in one rank on the WFD (Water Framework Directive) scale e.g. from clear to intermediate for one year. Further detail EvidenceBryozoans are suspension feeders that may be adversely affected by increases in suspended sediment, due to clogging of their feeding apparatus. However, Tyler-Walters & Ballerstedt (2007) reported Flustra foliacea as tolerant to increased suspended sediment based on its occurrence in areas of high suspended sediment e.g. abundant in turbid, fast flowing waters of the Menai Straits (Moore 1977). Communities dominated by Flustra foliacea were described on tide swept seabed, exposed to high levels of suspended sediment and sediment scour in the English Channel (Holme & Wilson 1985). Flustra foliacea is also characteristic of sediment-scoured, silty rock communities CR.HCR.XFa.FluCoAs and CR.MCR.EcCr.UrtScr (Connor et al., 2004). Despite sediment being generally considered to have a negative impact on suspension feeders (Gerrodette & Flechsig 1979), many encrusting sponges appear to be able survive in highly sedimented conditions, and many species prefer such habitats (Bell & Barnes 2001; Bell & Smith 2004). Castric & Chassé (1991) conducted a factorial analysis of the subtidal rocky ecology near Brest, France and rated the distribution of species in varying turbidity (corroborated by the depth at which laminarians disappeared). Cliona celata and Stelligera rigida were classed as indifferent to turbidity, Tethya aurantium, Pachymatisma johnstonia and Polymastia boletiformis (as Polymastia robusta) had a slight preference for clearer water, while Dysidea fragilis, Polymastia mamillaris, and Raspailia ramosa had a strong preference for turbid water. Storr (1976) observed the sponge Sphecispongia vesparium back washing to eject sediment and noted that other sponges (such as Condrilla nucula) use secretions to remove settled material. Raspailia ramosa and Stelligera stuposa have a reduced maximum size in areas of high sedimentation (Bell et al, 2002). Tjensvoll (2013) found that Geodia barretti physiologically shuts down when exposed to sediment concentrations of 100 mg /l (86% reduction). Rapid recovery to initial respiration levels directly after the exposure indicated that Geodia barretti can cope with a single short exposure to elevated sediment concentrations. However, it should be noted that a laboratory study on the impact of elevated sedimentation rates on deep water sponges found that sediment load of 30 mg sed./l resulted in significantly higher sponge mortality compared with sponges exposed to 5 and 10 mg sed./l, although no additional information was provided (Hoffman & Tore Rapp, pers com cited in Lancaster et al., 2014). Schönberg (2015) reviewed and observed the interactions between sediments and marine sponges and described the lack of research on Porifera, with most studies grouping them together when looking at sediment effects. Whilst many sponges are disadvantaged by sedimentation (as would be expected, being sessile filter feeders), many examples exist of sponges adapting to sediment presence, including through sediment incorporation, sediment encrusting, soft sediment anchoring using spicules and living, at least partially, embedded within the sediment. Among the characterizing species, Schönberg (2015) found that Polymastiida interacted with sediment in 18.9% of observations (primarily through spicules), Clionaida had a highly variable interaction with sediment, with 5.7±11.4 %, Tethyida interacted in 13.1±21.1%. Sensitivity assessment. The CR.HCR.XFa.ByErSp complex tends to occur in areas of high energy, therefore an increase in suspended sediment will result in an increase in scour. While some sponges and Flustra are probably resistant of scour, other fragile sponges, and bryozoans will be removed. The biotope will probably come to resemble Flustra dominated or grazed faunal turf communities in the short-term, and the biotope will be lost. Therefore, a resistance of ‘Low’ is suggested, with resilience of ‘Medium’ and sensitivity of ‘Medium’. | LowHelp | MediumHelp | MediumHelp |
Smothering and siltation rate changes (light) [Show more]Smothering and siltation rate changes (light)Benchmark. ‘Light’ deposition of up to 5 cm of fine material added to the seabed in a single discrete event. Further detail EvidenceSmothering by 5 cm of sediment is likely to prevent feeding, and hence growth and reproduction, as well as respiration in the bryozoans. In addition, associated sediment abrasion may remove the bryozoan colonies. A layer of sediment will probably also interfere with larval settlement (Tyler-Walters, 2005). Despite sediment being generally considered to have a negative impact on suspension feeders (Gerrodette & Flechsig 1979), many encrusting sponges appear to be able survive in highly sedimented conditions, and in fact many species prefer such habitats (Bell & Barnes 2001; Bell & Smith 2004). However, Wulff (2006) described mortality in three sponge groups following four weeks of burial under sediment. 16% of Amphimedon biomass died compared with 40% and 47% in Iotrochota and Aplysina respectively. The complete disappearance of the sea squirt Ascidiella aspera biocoenosis and associated sponges in the Black Sea near the Kerch Strait was attributed to siltation (Terent'ev, 2008 cited in Tillin & Tyler-Walters, 2014). Holme & Wilson (1985) examined the fauna in a tide-swept region of the central English Channel. Flustra foliacea dominated communities subject to sediment transport (mainly sand) and periodic, temporary, burial (ca <5 cm). Sensitivity assessment. Smothering by 5 cm of sediment is likely to cause limited mortality amongst some of characterizing species of this biotope (particularly the smaller sponges). This biotope tends to occur in areas with moderate to strong water movement and removal of the sediment is likely to be rapid. Therefore, resistance has been assessed as ‘High’. Resilience has been assessed as ‘High’ and sensitivity has been assessed as ‘Not sensitive’ at the benchmark level. | HighHelp | HighHelp | Not sensitiveHelp |
Smothering and siltation rate changes (heavy) [Show more]Smothering and siltation rate changes (heavy)Benchmark. ‘Heavy’ deposition of up to 30 cm of fine material added to the seabed in a single discrete event. Further detail EvidenceSmothering by 30 cm of sediment is likely to prevent feeding, and hence growth and reproduction, as well as respiration in the bryozoans. In addition, associated sediment abrasion may remove the bryozoan colonies. Sediment will probably also interfere with larval settlement (Tyler-Walters, 2005). Despite sediment being generally considered to have a negative impact on suspension feeders (Gerrodette & Flechsig 1979), many encrusting sponges appear to be able survive in highly sedimented conditions, and in fact many species prefer such habitats (Bell & Barnes 2001; Bell & Smith 2004). However, Wulff (2006) described mortality in three sponge groups following four weeks of burial under sediment. 16% of Amphimedon biomass died compared with 40% and 47% in Iotrochota and Aplysina respectively. The complete disappearance of the sea squirt Ascidiella aspera biocoenosis and associated sponges in the Black Sea near the Kerch Strait was attributed to siltation (Terent'ev 2008 cited in Tillin & Tyler-Walters, 2014). The majority of the characterizing species are likely to be buried in 30cm of sediment deposition. Sensitivity assessment. Smothering by 30 cm of sediment could cause mortality amongst the majority of characterizing species of this biotope, if it remained. However, this biotope occurs in areas with moderate to strong water movement and the deposited sediment will be removed quickly. Vertical surfaces may also afford protection, so that the effects will depend on the topography of the substratum. Resistance at the benchmark has been assessed as ‘Medium’. Resilience has been assessed as ‘High’, assuming sediment removal. Sensitivity has been assessed as ‘Low’. | MediumHelp | HighHelp | LowHelp |
Litter [Show more]LitterBenchmark. The introduction of man-made objects able to cause physical harm (surface, water column, seafloor or strandline). Further detail EvidenceNot assessed. | Not Assessed (NA)Help | Not assessed (NA)Help | Not assessed (NA)Help |
Electromagnetic changes [Show more]Electromagnetic changesBenchmark. A local electric field of 1 V/m or a local magnetic field of 10 µT. Further detail Evidence'No evidence'. | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Underwater noise changes [Show more]Underwater noise changesBenchmark. MSFD indicator levels (SEL or peak SPL) exceeded for 20% of days in a calendar year. Further detail EvidenceStanley et al. (2014) studied the effects of vessel noise on fouling communities and found that the bryozoans Bugula neritina, Watersipora arcuate and Watersipora subtorquata responded positively. More than twice as many bryozoans settled and established on surfaces with vessel noise (128 dB in the 30–10,000 Hz range) compared to those in silent conditions. Growth was also significantly higher in bryozoans exposed to noise, with 20% higher growth rate in encrusting and 35% higher growth rate in branching species. Sensitivity assessment. Whilst no evidence could be found for the effect of noise or vibrations on the characterizing sponges, it is unlikely that these species would be adversely affected by noise. Resistance to this pressure is assessed as 'High' and resilience as 'High'. This biotope is therefore considered to be 'Not sensitive'. | HighHelp | HighHelp | Not sensitiveHelp |
Introduction of light or shading [Show more]Introduction of light or shadingBenchmark. A change in incident light via anthropogenic means. Further detail EvidenceJones et al. (2012) found that many sponges, around Skomer Island, particularly encrusting species, preferred vertical or shaded bedrock to open, light surfaces, presumably due to lack of competition from algae. Sensitivity assessment Whilst sponges seem to favour shaded areas in which to settle, it is unlikely that changes at the benchmark pressure would result in mortality. Resistance to this pressure is assessed as 'High' and resilience as 'High'. This biotope is therefore considered to be 'Not sensitive'. | HighHelp | HighHelp | Not sensitiveHelp |
Barrier to species movement [Show more]Barrier to species movementBenchmark. A permanent or temporary barrier to species movement over ≥50% of water body width or a 10% change in tidal excursion. Further detail Evidence'Not relevant' as barriers and changes in tidal excursion are not relevant to biotopes restricted to open waters. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Death or injury by collision [Show more]Death or injury by collisionBenchmark. Injury or mortality from collisions of biota with both static or moving structures due to 0.1% of tidal volume on an average tide, passing through an artificial structure. Further detail Evidence'Not relevant' to seabed habitats. NB. Collision by grounding vessels is addressed under ‘surface abrasion’. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Visual disturbance [Show more]Visual disturbanceBenchmark. The daily duration of transient visual cues exceeds 10% of the period of site occupancy by the feature. Further detail Evidence'Not relevant' | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Biological Pressures
Use [show more] / [show less] to open/close text displayed
| Resistance | Resilience | Sensitivity | |
Genetic modification & translocation of indigenous species [Show more]Genetic modification & translocation of indigenous speciesBenchmark. Translocation of indigenous species or the introduction of genetically modified or genetically different populations of indigenous species that may result in changes in the genetic structure of local populations, hybridization, or change in community structure. Further detail EvidenceBugulina spp. are classed as fouling bryozoans and may be found in the intake pipes of ships or power stations, and on ship's hulls. The geographic distribution of Bugula species has been extended by transportation by shipping (Ryland, 1967). However, no information on the translocation of Bugula turbinata was found. In addition, no evidence of genetic modification or hybridization was found. Therefore, there is currently ‘No evidence’ on which to assess this pressure. | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Introduction of microbial pathogens [Show more]Introduction of microbial pathogensBenchmark. The introduction of relevant microbial pathogens or metazoan disease vectors to an area where they are currently not present (e.g. Martelia refringens and Bonamia, Avian influenza virus, viral Haemorrhagic Septicaemia virus). Further detail EvidenceGochfeld et al. (2012) found that diseased sponges hosted significantly different bacterial assemblages compared to healthy sponges, with diseased sponges also exhibiting significant decline in sponge mass and protein content. Sponge disease epidemics can have serious long-term effects on sponge populations, especially in long-lived, slow-growing species (Webster, 2007). Numerous sponge populations have been brought to the brink of extinction, including cases in the Caribbean with 70-95% disappearance of sponge specimens (Galstoff,1942),and in the Mediterranean (Vacelet,1994 cited in Cebrian et al., 2011; Gaino et al.,1992). Decaying patches and white bacterial film were reported in Haliclona oculata and Halichondria panicea in North Wales, 1988-89, (Webster, 2007). Specimens of Cliona spp. have exhibited blackened damage since 2013 in Skomer. Preliminary results have shown that clean, fouled and blackened Cliona all have very different bacterial communities. The blackened Cliona were effectively dead and had a bacterial community similar to marine sediments. The fouled Cliona had a very distinct bacterial community which may suggest a specific pathogen caused the effect (Burton, pers. comm..; Preston & Burton, 2015). No evidence for disease in the characterizing bryozoans could be found. Sensitivity assessment Sponge diseases have caused limited mortality in some species in the British Isles, although mass mortality and even extinction have been reported further afield. Whilst research is on-going into sponge disease in the UK, there is ‘No evidence’ to support an assessment of mortality due to diseasing in the characterizing species of this biotope. | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Removal of target species [Show more]Removal of target speciesBenchmark. Removal of species targeted by fishery, shellfishery or harvesting at a commercial or recreational scale. Further detail EvidenceSpongia officinalis (a Mediterranean species) has been targeted as a commercial species for use as bath sponges, although this species does not occur in the British Isles and no record of commercial exploitation of sponges in the British Isles could be found. No evidence for commercial exploitation of bryozoans could be found. Should removal of target species occur, the sessile, epifaunal nature of the characterizing species would result in little resistance to this pressure. This pressure is ‘Not relevant’ as none of the characterizing species are targeted. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Removal of non-target species [Show more]Removal of non-target speciesBenchmark. Removal of features or incidental non-targeted catch (by-catch) through targeted fishery, shellfishery or harvesting at a commercial or recreational scale. Further detail EvidenceThe characteristic species probably compete for space within the biotope, so that loss of one species would probably have little if any effect on the other members of the community. However, removal of the characteristic epifauna due to by-catch is likely to remove a proportion of the biotope and change the biological character of the biotope. These direct, physical impacts are assessed through the abrasion and penetration of the seabed pressures. The sensitivity assessment for this pressure considers any biological/ecological effects resulting from the removal of non-target species on this biotope. Whilst a large proportion of the sponge community is likely to be affected by abrasion events, there is some debate as it the level of effects depending on the size of the sponge and the type of abrasion effect (see Freese et al., 1999 and Coleman et al., 2013). However, the incidental removal of a proportion of the sponge and bryozoans populations, would result in a change in the character of the biotope. Therefore, resistance is recorded as ‘Low’, resilience is recorded as ‘Medium’ and sensitivity is ‘Medium’. | LowHelp | MediumHelp | MediumHelp |
Introduction or spread of invasive non-indigenous species (INIS) Pressures
Use [show more] / [show less] to open/close text displayed
| Resistance | Resilience | Sensitivity | |
Other INIS [Show more]Other INISEvidenceThis biotope is classified as circalittoral and therefore no algal species have been considered. Crepidula fornicata larvae require hard substrata for settlement. It prefers muddy gravelly, shell-rich, substrata that include gravel, or shells of other Crepidula, or other species e.g., oysters, and mussels. It is highly gregarious and seeks out adult shells for settlement, forming characteristic ‘stacks’ of adults. But it also recorded from rock, artificial substrata, and Sabellaria alveolata reefs (Blanchard, 1997, 2009; Bohn et al., 2012, 2013a, 2013b, 2015; De Montaudouin et al., 2018; Hinz et al., 2011; Helmer et al., 2019; Powell-Jennings & Calloway, 2018; Preston et al., 2020; Tillin et al., 2020). Close examination of the literature (2023) shows that evidence of its colonization and density on bedrock in the infralittoral or circalittoral was lacking. Tillin et al. (2020) suggested that Crepidula could colonize circalittoral rock due to its presence on tide-swept rough grounds in the English Channel (Hinz et al., 2011). However, Hinz et al. (2011) reported that Crepidula fornicata only dominated one assemblage (with an average of 181 individuals per trawl) on gravel substratum with boulders. Bohn et al. (2015) noted that Crepidula occurred at low density or was absent in areas dominated by boulders, and Bohn et al. (2013a, 2013b, 2015) and Preston et al. (2020) showed that while Crepidula could settle on slate panels or ‘stone’ it preferred shell, especially that of conspecifics. In addition, no evidence was found of the effect of Crepidula populations on faunal turf-dominated habitats. It was only recorded at low density (0.1-0.9/m2) in one faunal turf biotope (CR.MCR.CFaVS.CuSpH.As) (JNCC, 2015). Faunal turfs are dominated by suspension feeders so larval predation is probably high, which may prevent colonization by Crepidula. Also, faunal turf species actively compete for space and many are fast growing and opportunistic, so may out-compete Crepidula for space even if it gained a foothold in the community. Didemnum vexillum is an invasive colonial sea squirt native to Asia which was first recorded in the UK in Darthaven Marina, Dartmouth in 2005. Didemnum vexillum can form extensive mats over the substrata it colonizes; binding boulders and cobbles and altering the host habitat (Griffith et al., 2009). Didemnum vexillum can also grow over and smother the resident biological community. Recent surveys within Holyhead Marina, North Wales have found Didemnum vexillum growing on and smothering native tunicate communities, including Ciona intestinalis (Griffith et al., 2009). Due to the rapid-re-colonization of Didemnum vexillum eradication attempts have to date failed. Presently Didemnum vexillum is isolated to several sheltered locations in the UK (NBN, 2015). However Didemnum vexillum has successfully colonized the offshore location of the Georges Bank, USA (Lengyel et al., 2009) which is more exposed than the locations which Didemnum vexillum have colonized in the UK. It is therefore possible that Didemnum vexillum could colonize more exposed locations within the UK and could therefore pose a threat to these biotopes. Several invasive bryozoans are of concern, including Schizoporella japonica (Ryland et al., 2014) and Tricellaria inopinata (Dyrynda et al., 2000; Cook et al., 2013), however, evidence of potential effects is sparse. Sensitivity assessment. The circalittoral rock characterizing this biotope is likely to be unsuitable for the colonization by Crepidula fornicata due to the extremely wave exposed to moderately wave exposed conditions, in which wave action and storms may mitigate or prevent the colonization by Crepidula at high densities, although Crepidula has been recorded from areas of strong tidal streams (Hinz et al., 2011). Crepidula has been recorded from the lower intertidal to ca 160 m in depth but is most common in the shallow subtidal above 50 m (Blanchard, 1997; Thieltges et al., 2003; Bohn et al., 2012, 2015; Hinz et al., 2011; OBIS, 2023; Tillin et al., 2020), therefore, colonization of Crepidula would be limited to low densities in deeper examples of the biotope. In addition, no evidence was found of the effect of Crepidula populations on faunal turf-dominated habitats or infralittoral or circalittoral rock habitats. At present, there is 'Insufficient evidence' to suggest that the circalittoral rock biotopes are sensitive to colonization by Crepidula fornicata or other invasive species; further evidence is required. | Insufficient evidence (IEv)Help | Not relevant (NR)Help | Help |
Bibliography
Ackers, R.G., 1983. Some local and national distributions of sponges. Porcupine Newsletter, 2 (7).
Ackers, R.G.A., Moss, D. & Picton, B.E. 1992. Sponges of the British Isles (Sponges: V): a colour guide and working document. Ross-on-Wye: Marine Conservation Society.
Ager, O.E.D. 2007. Alcyonidium diaphanum, Sea chervil. In Tyler-Walters H. and Hiscock K. (eds) Marine Life Information Network: Biology and Sensitivity Key Information Reviews, [on-line]. Plymouth: Marine Biological Association of the United Kingdom. Available from: http://www.marlin.ac.uk/species/detail/1738
Banks, P.D. & Brown, K.M., 2002. Hydrocarbon effects on fouling assemblages: the importance of taxonomic differences, seasonal, and tidal variation. Marine Environmental Research, 53 (3), 311-326.
Bell, J.J. & Barnes, D.K., 2000. The distribution and prevalence of sponges in relation to environmental gradients within a temperate sea lough: inclined cliff surfaces. Diversity and Distributions, 6 (6), 305-323.
Bell, J.J. & Barnes, D.K., 2001. Sponge morphological diversity: a qualitative predictor of species diversity? Aquatic Conservation: Marine and Freshwater Ecosystems, 11 (2), 109-121.
Bell, J.J. & Smith, D., 2004. Ecology of sponge assemblages (Porifera) in the Wakatobi region, south-east Sulawesi, Indonesia: richness and abundance. Journal of the Marine Biological Association of the UK, 84 (3), 581-591.
Bell, J.J., Barnes, D. & Shaw, C., 2002. Branching dynamics of two species of arborescent demosponge: the effect of flow regime and bathymetry. Journal of the Marine Biological Association of the UK, 82 (2), 279-294.
Berman, J., Burton, M., Gibbs, R., Lock, K., Newman, P., Jones, J. & Bell, J., 2013. Testing the suitability of a morphological monitoring approach for identifying temporal variability in a temperate sponge assemblage. Journal for Nature Conservation, 21 (3), 173-182.
Biggs, B.C., 2013. Harnessing Natural Recovery Processes to Improve Restoration Outcomes: An Experimental Assessment of Sponge-Mediated Coral Reef Restoration. Plos One, 8 (6), e64945.
Blanchard, M., 2009. Recent expansion of the slipper limpet population (Crepidula fornicata) in the Bay of Mont-Saint-Michel (Western Channel, France). Aquatic Living Resources, 22 (1), 11-19. DOI https://doi.org/10.1051/alr/2009004
Blanchard, M., 1997. Spread of the slipper limpet Crepidula fornicata (L.1758) in Europe. Current state and consequences. Scientia Marina, 61, Supplement 9, 109-118. Available from: http://scimar.icm.csic.es/scimar/index.php/secId/6/IdArt/290/
Bohn, K., Richardson, C. & Jenkins, S., 2012. The invasive gastropod Crepidula fornicata: reproduction and recruitment in the intertidal at its northernmost range in Wales, UK, and implications for its secondary spread. Marine Biology, 159 (9), 2091-2103. DOI https://doi.org/10.1007/s00227-012-1997-3
Bohn, K., Richardson, C.A. & Jenkins, S.R., 2015. The distribution of the invasive non-native gastropod Crepidula fornicata in the Milford Haven Waterway, its northernmost population along the west coast of Britain. Helgoland Marine Research, 69 (4), 313.
Bohn, K., Richardson, C.A. & Jenkins, S.R., 2013a. Larval microhabitat associations of the non-native gastropod Crepidula fornicata and effects on recruitment success in the intertidal zone. Journal of Experimental Marine Biology and Ecology, 448, 289-297. DOI https://doi.org/10.1016/j.jembe.2013.07.020
Bohn, K., Richardson, C.A. & Jenkins, S.R., 2013b. The importance of larval supply, larval habitat selection and post-settlement mortality in determining intertidal adult abundance of the invasive gastropod Crepidula fornicata. Journal of Experimental Marine Biology and Ecology, 440, 132-140. DOI https://doi.org/10.1016/j.jembe.2012.12.008
Boulcott, P. & Howell, T.R.W., 2011. The impact of scallop dredging on rocky-reef substrata. Fisheries Research (Amsterdam), 110 (3), 415-420.
Boury-Esnault, N., 1987. The Polymastia species (Demosponges, Hadromerida) of the Atlantic area. Taxonomy of Porifera: Springer, pp. 29-66.
Budd, G.C. 2008. Alcyonium digitatum Dead man's fingers. In Tyler-Walters H. and Hiscock K. (eds) Marine Life Information Network: Biology and Sensitivity Key Information Reviews, [on-line]. Plymouth: Marine Biological Association of the United Kingdom. Available from: http://www.marlin.ac.uk/species/detail/1187
Bullimore, B., 1985. An investigation into the effects of scallop dredging within the Skomer Marine Reserve. Report to the Nature Conservancy Council by the Skomer Marine Reserve Subtidal Monitoring Project, S.M.R.S.M.P. Report, no 3., Nature Conservancy Council.
Carver, C., Mallet, A. & Vercaemer, B., 2006. Biological synopsis of the solitary tunicate Ciona intestinalis. Canadian Manuscript Report of Fisheries and Aquatic Science, No. 2746, v + 55 p. Bedford Institute of Oceanography, Dartmouth, Nova Scotia.
Castège, I., Milon, E. & Pautrizel, F., 2014. Response of benthic macrofauna to an oil pollution: Lessons from the “Prestige” oil spill on the rocky shore of Guéthary (south of the Bay of Biscay, France). Deep Sea Research Part II: Topical Studies in Oceanography, 106, 192-197.
Castric-Fey, A. & Chassé, C., 1991. Factorial analysis in the ecology of rocky subtidal areas near Brest (west Brittany, France). Journal of the Marine Biological Association of the United Kingdom, 71, 515-536.
Cebrian, E., Uriz, M.J., Garrabou, J. & Ballesteros, E., 2011. Sponge mass mortalities in a warming Mediterranean Sea: are cyanobacteria-harboring species worse off? Plos One, 6 (6), e20211.
Chia, F.S. & Spaulding, J.G., 1972. Development and juvenile growth of the sea anemone Tealia crassicornis. Biological Bulletin, Marine Biological Laboratory, Woods Hole, 142, 206-218.
Cocito, S. & Sgorbini, S., 2014. Long-term trend in substratum occupation by a clonal, carbonate bryozoan in a temperate rocky reef in times of thermal anomalies. Marine Biology, 161 (1), 17-27.
Cocito, S., Ferdeghini, F., & Sgorbini, S., 1998b. Pentapora fascialis (Pallas) [Cheilostomata: Ascophora] colonization of one sublittoral rocky site after sea-storm in the northwest Mediterranean. Hydrobiologia, 375/376, 59-66.
Cole, S., Codling, I.D., Parr, W. & Zabel, T., 1999. Guidelines for managing water quality impacts within UK European Marine sites. Natura 2000 report prepared for the UK Marine SACs Project. 441 pp., Swindon: Water Research Council on behalf of EN, SNH, CCW, JNCC, SAMS and EHS. [UK Marine SACs Project.]. Available from: http://ukmpa.marinebiodiversity.org/uk_sacs/pdfs/water_quality.pdf
Coleman, R.A., Hoskin, M.G., von Carlshausen, E. & Davis, C.M., 2013. Using a no-take zone to assess the impacts of fishing: Sessile epifauna appear insensitive to environmental disturbances from commercial potting. Journal of Experimental Marine Biology and Ecology, 440, 100-107.
Connor, D.W., Allen, J.H., Golding, N., Howell, K.L., Lieberknecht, L.M., Northen, K.O. & Reker, J.B., 2004. The Marine Habitat Classification for Britain and Ireland. Version 04.05. ISBN 1 861 07561 8. In JNCC (2015), The Marine Habitat Classification for Britain and Ireland Version 15.03. [2019-07-24]. Joint Nature Conservation Committee, Peterborough. Available from https://mhc.jncc.gov.uk/
Cook, R., Fariñas-Franco, J. M., Gell, F. R., Holt, R. H., Holt, T., Lindenbaum, C., Porter, J.S., Seed, R., Skates, L.R., Stringell, T.B. & Sanderson, W.G., 2013. The substantial first impact of bottom fishing on rare biodiversity hotspots: a dilemma for evidence-based conservation. PloS One, 8 (8), e69904. DOI https://doi.org/10.1371/journal.pone.0069904
Crisp, D.J. (ed.), 1964. The effects of the severe winter of 1962-63 on marine life in Britain. Journal of Animal Ecology, 33, 165-210.
De Montaudouin, X., Blanchet, H. & Hippert, B., 2018. Relationship between the invasive slipper limpet Crepidula fornicata and benthic megafauna structure and diversity, in Arcachon Bay. Journal of the Marine Biological Association of the United Kingdom, 98 (8), 2017-2028. DOI https://doi.org/10.1017/s0025315417001655
De Vos, L., Rútzler K., Boury-Esnault, N., Donadey C., Vacelet, J., 1991. Atlas of Sponge Morphology. Atlas de Morphologie des Éponges. Washington, Smithsonian Institution Press.
Diaz, R.J. & Rosenberg, R., 1995. Marine benthic hypoxia: a review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanography and Marine Biology: an Annual Review, 33, 245-303.
Dyrynda, P.E.J. & Ryland, J.S., 1982. Reproductive strategies and life histories in the cheilostome marine bryozoans Chartella papyracea and Bugula flabellata. Marine Biology, 71, 241-256.
Dyrynda, P.E.J., 1994. Hydrodynamic gradients and bryozoan distributions within an estuarine basin (Poole Harbour, UK). In Proceedings of the 9th International Bryozoology conference, Swansea, 1992. Biology and Palaeobiology of Bryozoans (ed. P.J. Hayward, J.S. Ryland & P.D. Taylor), pp.57-63. Fredensborg: Olsen & Olsen.
Eggleston, D., 1972a. Patterns of reproduction in marine Ectoprocta off the Isle of Man. Journal of Natural History, 6, 31-38.
Eggleston, D., 1972b. Factors influencing the distribution of sub-littoral ectoprocts off the south of the Isle of Man (Irish Sea). Journal of Natural History, 6, 247-260.
Eno, N.C., Clark, R.A. & Sanderson, W.G. (ed.) 1997. Non-native marine species in British waters: a review and directory. Peterborough: Joint Nature Conservation Committee.
Fehlauer-Ale, K.H., Winston, J.E., Tilbrook, K.J., Nascimento, K.B. & Vieira, L.M., 2015. Identifying monophyletic groups within Bugula sensu lato (Bryozoa, Buguloidea). Zoologica Scripta, 44 (3), 334-347.
Fish, J.D. & Fish, S., 1996. A student's guide to the seashore. Cambridge: Cambridge University Press.
Fortunato, H., Schäfer, P. & Blaschek, H., 2013. Growth Rates, Age Determination, and Calcification Levels in Flustra foliacea (L.) (Bryozoa: Cheilostomata): Preliminary Assessment. In Ernst, A., et al. (eds.). Bryozoan Studies 2010, Berlin, Heidelberg: Springer Berlin Heidelberg, pp. 59-74.
Fowler, S. & Laffoley, D., 1993. Stability in Mediterranean-Atlantic sessile epifaunal communities at the northern limits of their range. Journal of Experimental Marine Biology and Ecology, 172 (1), 109-127. DOI https://doi.org/10.1016/0022-0981(93)90092-3
Freese, J.L., 2001. Trawl-induced damage to sponges observed from a research submersible. Marine Fisheries Review, 63 (3), 7-13.
Freese, L., Auster, P.J., Heifetz, J. & Wing, B.L., 1999. Effects of trawling on seafloor habitat and associated invertebrate taxa in the Gulf of Alaska. Marine Ecology Progress Series, 182, 119-126.
Gaino, E., Pronzato, R., Corriero, G. & Buffa, P., 1992. Mortality of commercial sponges: incidence in two Mediterranean areas. Italian Journal of Zoology, 59 (1), 79-85.
Galstoff, P., 1942. Wasting disease causing mortality of sponges in the West Indies and Gulf of Mexico. Proceedings 8th American Scientific Congress, pp. 411-421.
Galtsoff, P., 1964. The American Oyster Crassostrea virginica Gmelin. Fishery Bulletin of the Fish and Wildlife Service, 64, 1–480
Gerrodette, T. & Flechsig, A., 1979. Sediment-induced reduction in the pumping rate of the tropical sponge Verongia lacunosa. Marine Biology, 55 (2), 103-110.
Gochfeld, D., Easson, C., Freeman, C., Thacker, R. & Olson, J., 2012. Disease and nutrient enrichment as potential stressors on the Caribbean sponge Aplysina cauliformis and its bacterial symbionts. Marine Ecology Progress Series, 456, 101-111.
Gochfeld, D.J., Schlöder, C. & Thacker, R.W., 2007. Sponge community structure and disease prevalence on coral reefs in Bocas del Toro, Panama. Porifera Research: Biodiversity, Innovation, and Sustainability, Série Livros, 28, 335-343.
Griffith, K., Mowat, S., Holt, R.H., Ramsay, K., Bishop, J.D., Lambert, G. & Jenkins, S.R., 2009. First records in Great Britain of the invasive colonial ascidian Didemnum vexillum Kott, 2002. Aquatic Invasions, 4 (4), 581-590.
Gunda, V.G. & Janapala, V.R., 2009. Effects of dissolved oxygen levels on survival and growth in vitro of Haliclona pigmentifera (Demospongiae). Cell and tissue research, 337 (3), 527-535.
Hartikainen, H., Johnes, P., Moncrieff, C. & Okamura, B., 2009. Bryozoan populations reflect nutrient enrichment and productivity gradients in rivers. Freshwater Biology, 54 (11), 2320-2334.
Hayward, P.J. & Ryland, J.S. 1979. British ascophoran bryozoans. London: Academic Press.
Hayward, P.J. & Ryland, J.S. 1994. The marine fauna of the British Isles and north-west Europe. Volume 1. Introduction and Protozoans to Arthropods. Oxford: Clarendon Press.
Hayward, P.J. & Ryland, J.S. 1998. Cheilostomatous Bryozoa. Part 1. Aeteoidea - Cribrilinoidea. Shrewsbury: Field Studies Council. [Synopses of the British Fauna, no. 10. (2nd edition)]
Hayward, P.J. & Ryland, J.S. (ed.), 1995. The marine fauna of the British Isles and north-west Europe. Volume 2. Molluscs to Chordates. Oxford Science Publications. Oxford: Clarendon Press.
Helmer, L., Farrell, P., Hendy, I., Harding, S., Robertson, M. & Preston, J., 2019. Active management is required to turn the tide for depleted Ostrea edulis stocks from the effects of overfishing, disease and invasive species. Peerj, 7 (2). DOI https://doi.org/10.7717/peerj.6431
Herreid, C.F., 1980. Hypoxia in invertebrates. Comparative Biochemistry and Physiology Part A: Physiology, 67 (3), 311-320. DOI https://doi.org/10.1016/S0300-9629(80)80002-8
Hincks, T., 1880. A history of British marine Polyzoa, vol. I & II. London: John van Voorst.
Hinz, H., Capasso, E., Lilley, M., Frost, M. & Jenkins, S.R., 2011. Temporal differences across a bio-geographical boundary reveal slow response of sub-littoral benthos to climate change. Marine Ecology Progress Series, 423, 69-82. DOI https://doi.org/10.3354/meps08963
Hiscock, K. & Hoare, R., 1975. The ecology of sublittoral communities at Abereiddy Quarry, Pembrokeshire. Journal of the Marine Biological Association of the United Kingdom, 55 (4), 833-864.
Hiscock, K., 1981. Marine life on the wreck of the M.V. "Robert". Report of the Lundy Field Society, 32, 40-44.
Hiscock, K., 1983. Water movement. In Sublittoral ecology. The ecology of shallow sublittoral benthos (ed. R. Earll & D.G. Erwin), pp. 58-96. Oxford: Clarendon Press.
Hiscock, K., 1985. Littoral and sublittoral monitoring in the Isles of Scilly. September 22nd to 29th, 1984. Nature Conservancy Council, Peterborough, CSD Report, no. 562., Field Studies Council Oil Pollution Research Unit, Pembroke.
Hiscock, K., 1994. Marine communities at Lundy - origins, longevity and change. Biological Journal of the Linnean Society 51, 183-188.
Hiscock, K., 2002. Changes in the marine life of Lundy. Report of the Lundy Field Society. 52, 84-93. Available from https://lfs-resources.s3.amazonaws.com/ar52/LFS_Annual_Report_Vol_52_Part_16.pdf
Holme, N.A. & Wilson, J.B., 1985. Faunas associated with longitudinal furrows and sand ribbons in a tide-swept area in the English Channel. Journal of the Marine Biological Association of the United Kingdom, 65, 1051-1072.
Jackson, A., 2008. Ciona intestinalis. A sea squirt. Marine Life Information Network: Biology and Sensitivity Key Information Sub-programme [On-line]. Plymouth: Marine Biological Association of the United Kingdom. [cited 16/12/15] Available from:
Jackson, A. 2016. Pentapora foliacea (Ross). In Tyler-Walters H. and Hiscock K. (eds) Marine Life Information Network: Biology and Sensitivity Key Information Reviews, [on-line]. Plymouth: Marine Biological Association of the United Kingdom. [cited Available from: http://www.marlin.ac.uk/species/detail/1389
Jackson, E.L., Langmead, O., Barnes, M., Tyler-Walters, H. & Hiscock, K., 2008. Identification of indicator species to represent the full range of benthic life history strategies for Lyme Bay and the consideration of the wider application for monitoring of Marine Protected Areas. Report to the Department of Environment, Food and Rural Affairs from the Marine Life Information Network (MarLIN). Plymouth: Marine Biological Association of the UK. Defra Contract No. MB101 Milestone 2.
Jennings, S. & Kaiser, M.J., 1998. The effects of fishing on marine ecosystems. Advances in Marine Biology, 34, 201-352.
JNCC (Joint Nature Conservation Committee), 2022. The Marine Habitat Classification for Britain and Ireland Version 22.04. [Date accessed]. Available from: https://mhc.jncc.gov.uk/
Jones, J., Bunker, F., Newman, P., Burton, M., Lock, K., 2012. Sponge Diversity of Skomer Marine Nature Reserve. CCW Regional Report, CCW/WW/12/3.
Knight-Jones, E.W. & Nelson-Smith, A., 1977. Sublittoral transects in the Menai Straits and Milford Haven. In Biology of benthic organisms (ed. B.F. Keegan, P. O Ceidigh & P.J.S. Broaden), pp. 379-390. Oxford: Pergamon Press.
Lancaster, J. (ed), McCallum, S., A.C., L., Taylor, E., A., C. & Pomfret, J., 2014. Development of Detailed Ecological Guidance to Support the Application of the Scottish MPA Selection Guidelines in Scotland’s seas. Scottish Natural Heritage Commissioned Report No.491 (29245), Scottish Natural Heritage, Inverness, 40 pp.
Lengyel, N.L., Collie, J.S. & Valentine, P.C., 2009. The invasive colonial ascidian Didemnum vexillum on Georges Bank - Ecological effects and genetic identification. Aquatic Invasions, 4(1), 143-152. DOI https://doi.org/10.3391/ai.2009.4.1.15
Lewis, J.R., 1964. The Ecology of Rocky Shores. London: English Universities Press.
Lock, K., Burton, M., Luddington, L. & Newman, P., 2006. Skomer Marine Nature Reserve project status report 2005/06. Countryside Council for Wales, Bangor, CCW Regional Report CCW/WW/05/9.
Lombardi, C., Taylor, P.D. & Cocito, S., 2010. Systematics of the Miocene–Recent bryozoan genus Pentapora (Cheilostomata). Zoological Journal of the Linnean Society, 160 (1), 17-39. DOI: 10.1111/j.1096-3642.2009.00594.x
Manuel, R.L., 1988. British Anthozoa. Synopses of the British Fauna (New Series) (ed. D.M. Kermack & R.S.K. Barnes). The Linnean Society of London [Synopses of the British Fauna No. 18.]. DOI https://doi.org/10.1002/iroh.19810660505
Marin, A., Lopez, M., Esteban, M., Meseguer, J., Munoz, J. & Fontana, A., 1998. Anatomical and ultrastructural studies of chemical defence in the sponge Dysidea fragilis. Marine Biology, 131 (4), 639-645.
Mazouni, N., Gaertner, J. & Deslous-Paoli, J.-M., 2001. Composition of biofouling communities on suspended oyster cultures: an in situ study of their interactions with the water column. Marine Ecology Progress Series, 214, 93-102.
McKinney, F.K., 1986. Evolution of erect marine bryozoan faunas: repeated success of unilaminate species The American Naturalist, 128, 795-809.
Moore, P.G., 1977a. Inorganic particulate suspensions in the sea and their effects on marine animals. Oceanography and Marine Biology: An Annual Review, 15, 225-363.
Naylor. P., 2011. Great British Marine Animals, 3rd Edition. Plymouth. Sound Diving Publications.
NBN, 2015. National Biodiversity Network 2015(20/05/2015). https://data.nbn.org.uk/
O'Dea, A. & Okamura, B., 2000. Life history and environmental inference through retrospective morphometric analysis of bryozoans: a preliminary study. Journal of the Marine Biological Association of the United Kingdom, 80, 1127-1128.
OBIS (Ocean Biodiversity Information System), 2025. Global map of species distribution using gridded data. Available from: Ocean Biogeographic Information System. www.iobis.org. Accessed: 2025-07-15
Okamura, B., 1984. The effects of ambient flow velocity, colony size and upstream colonies on the feeding success of Bryozoa, Bugula stolonifera Ryland, an arborescent species. Journal of the Experimental Marine Biology and Ecology, 83, 179-193.
Patzold, J., Ristedt, H. & Wefer, G., 1987. Rate of growth and longevity of a large colony of Pentapora foliacea (Bryozoa) recorded in their oxygen isotope profiles. Marine Biology, 96, 535-538.
Picton, B.E. & Morrow, C.C., 2015. Ascidia mentula O F Müller, 1776. In Encyclopedia of Marine Life of Britain and Ireland. [cited 26/01/16]. Available from: http://www.habitas.org.uk/marinelife/species.asp?item=ZD1500
Powell-Jennings, C. & Callaway, R., 2018. The invasive, non-native slipper limpet Crepidula fornicata is poorly adapted to sediment burial. Marine Pollution Bulletin, 130, 95-104. DOI https://doi.org/10.1016/j.marpolbul.2018.03.006
Preston J. & Burton, M., 2015. Marine microbial assemblages associated with diseased Porifera in Skomer Marine Nature Reserve (SMNR), Wales. Aquatic Biodiversity and Ecosystems, 30th August – 4th September, Liverpool., pp. p110.
Preston, J., Fabra, M., Helmer, L., Johnson, E., Harris-Scott, E. & Hendy, I.W., 2020. Interactions of larval dynamics and substrate preference have ecological significance for benthic biodiversity and Ostrea edulis Linnaeus, 1758 in the presence of Crepidula fornicata. Aquatic Conservation: Marine and Freshwater Ecosystems, 30 (11), 2133-2149. DOI https://doi.org/10.1002/aqc.3446
Ramos, M., 2010. IBERFAUNA. The Iberian Fauna Databank, 2015(2015/12/21). http://iberfauna.mncn.csic.es/
Riisgård, H.U., Lüskow, F., Pleissner, D., Lundgreen, K. & López, M., 2013. Effect of salinity on filtration rates of mussels Mytilus edulis with special emphasis on dwarfed mussels from the low-saline Central Baltic Sea. Helgoland Marine Research, 67, 591-8
Roberts, D., Cummins, S., Davis, A. & Chapman, M., 2006. Structure and dynamics of sponge-dominated assemblages on exposed and sheltered temperate reefs. Marine Ecology Progress Series, 321, 19-30.
Rose, C.S. & Risk, M.J., 1985. Increase in Cliona delitrix infestation of Montastrea cavernosa heads on an organically polluted portion of the Grand Cayman fringing reef. Marine Ecology, 6 (4), 345-363.
Rosenberg, R., Hellman, B. & Johansson, B., 1991. Hypoxic tolerance of marine benthic fauna. Marine Ecology Progress Series, 79, 127-131. DOI https://dx.doi.org/10.3354/meps079127
Rosso, A., 2003. Bryozoan diversity in the Mediterranean Sea. Biogeographia, 24, 227-250.
Ryland, J.S., 1967. Polyzoa. Oceanography and Marine Biology: an Annual Review, 5, 343-369.
Ryland, J.S., 1970. Bryozoans. London: Hutchinson University Library.
Ryland, J.S., 1976. Physiology and ecology of marine bryozoans. Advances in Marine Biology, 14, 285-443.
Ryland, J.S., Holt, R., Loxton, J., Spencer Jones, M. & Porter, J.S., 2014. First occurrence of the non-native bryozoan Schizoporella japonica Ortmann (1890) in Western Europe. Zootaxa, 3780 (3), 481-502.
Sagasti, A., Schaffner, L.C. & Duffy, J.E., 2000. Epifaunal communities thrive in an estuary with hypoxic episodes. Estuaries, 23 (4), 474-487.
Schönberg, C.H.L., 2015. Happy relationships between marine sponges and sediments–a review and some observations from Australia. Journal of the Marine Biological Association of the United Kingdom, 1-22.
Sebens, K.P., 1981. Recruitment in a Sea Anemone Population: Juvenile Substrate Becomes Adult Prey. Science, 213 (4509), 785-787.
Sebens, K.P., 1985. Community ecology of vertical rock walls in the Gulf of Maine: small-scale processes and alternative community states. In The Ecology of Rocky Coasts: essays presented to J.R. Lewis, D.Sc. (ed. P.G. Moore & R. Seed), pp. 346-371. London: Hodder & Stoughton Ltd.
Sebens, K.P., 1986. Spatial relationships among encrusting marine organisms in the New England subtidal zone. Ecological Monographs, 56, 73-96. DOI https://doi.org/10.2307/2937271
Silén, L., 1981. Colony structure in Flustra foliacea (Linnaeus) (Bryozoa, Cheilostomata). Acta Zoologica (Stockholm.), 62, 219-232.
Soule, D.F. & Soule, J.D., 1979. Bryozoa (Ectoprocta). In Hart, C.W. & Fuller, S.L.H. (eds), Pollution ecology of estuarine invertebrates. New York: Academic Press, pp. 35-76.
Stanley, J.A., Wilkens, S.L. & Jeffs, A.G., 2014. Fouling in your own nest: vessel noise increases biofouling. Biofouling, 30 (7), 837-844.
Stebbing, A.R.D., 1971a. Growth of Flustra foliacea (Bryozoa). Marine Biology, 9, 267-273.
Storr, J.F. 1976. Ecological factors controlling sponge distribution in the Gulf of Mexico and the resulting zonation. In Aspects of Sponge Biology (ed. F.W. Harrison & R.R. Cowden), pp. 261-276. New York: Academic Press.
Thieltges, D.W., Strasser, M. & Reise, K., 2003. The American slipper-limpet Crepidula fornicata (L.) in the Northern Wadden Sea 70 years after its introduction. Helgoland Marine Research, 57, 27-33
Tillin, H. & Tyler-Walters, H., 2014b. Assessing the sensitivity of subtidal sedimentary habitats to pressures associated with marine activities. Phase 2 Report – Literature review and sensitivity assessments for ecological groups for circalittoral and offshore Level 5 biotopes. JNCC Report No. 512B, 260 pp. Available from: www.marlin.ac.uk/publications
Tillin, H.M., Kessel, C., Sewell, J., Wood, C.A. & Bishop, J.D.D., 2020. Assessing the impact of key Marine Invasive Non-Native Species on Welsh MPA habitat features, fisheries and aquaculture. NRW Evidence Report. Report No: 454. Natural Resources Wales, Bangor, 260 pp. Available from https://naturalresourceswales.gov.uk/media/696519/assessing-the-impact-of-key-marine-invasive-non-native-species-on-welsh-mpa-habitat-features-fisheries-and-aquaculture.pdf
Tilmant, J.T., 1979. Observations on the impact of shrimp roller frame trawls operated over hard-bottom communities, Biscayne Bay, Florida: National Park Service.
Tjensvoll, I., Kutti, T., Fosså, J.H. & Bannister, R., 2013. Rapid respiratory responses of the deep-water sponge Geodia barretti exposed to suspended sediments. Aquatic Biology, 19, 65-73.
Tyler-Walters, H., 2005. Laminaria hyperborea with dense foliose red seaweeds on exposed infralittoral rock. Marine Life Information Network: Biology and Sensitivity Key Information Sub-programme [on-line]: Plymouth: Marine Biological Association of the United Kingdom. 2015(20/05/2015). http://www.marlin.ac.uk/habitatsbasicinfo.php?habitatid=171&code=1997
Tyler-Walters, H. & Ballerstedt, S., 2007. Flustra foliacea Hornwrack. In Tyler-Walters H. and Hiscock K. (eds) Marine Life Information Network: Biology and Sensitivity Key Information Reviews, [on-line]. Plymouth: Marine Biological Association of the United Kingdom. Available from: http://www.marlin.ac.uk/species/detail/1609
Van Dolah, R.F., Wendt, P.H. & Nicholson, N., 1987. Effects of a research trawl on a hard-bottom assemblage of sponges and corals. Fisheries Research, 5 (1), 39-54.
Van Soest, R.W.M., Picton, B. & Morrow, C., 2000. Sponges of the North East Atlantic. [CD-ROM] Windows version 1.0. Amsterdam: Biodiversity Center of ETI, Multimedia Interactive Software. [World Biodiversity Database CD-ROM Series.]
Vethaak, A.D., Cronie, R.J.A. & van Soest, R.W.M., 1982. Ecology and distribution of two sympatric, closely related sponge species, Halichondria panicea (Pallas, 1766) and H. bowerbanki Burton, 1930 (Porifera, Demospongiae), with remarks on their speciation. Bijdragen tot de Dierkunde, 52, 82-102.
Ward-Paige, C.A., Risk, M.J., Sherwood, O.A. & Jaap, W.C., 2005. Clionid sponge surveys on the Florida Reef Tract suggest land-based nutrient inputs. Marine Pollution Bulletin, 51 (5), 570-579.
Webster, N.S., 2007. Sponge disease: a global threat? Environmental Microbiology, 9 (6), 1363-1375.
Webster, N.S. & Taylor, M.W., 2012. Marine sponges and their microbial symbionts: love and other relationships. Environmental Microbiology, 14 (2), 335-346.
Webster, N.S., Botté, E.S., Soo, R.M. & Whalan, S., 2011. The larval sponge holobiont exhibits high thermal tolerance. Environmental Microbiology Reports, 3 (6), 756-762.
Webster, N.S., Cobb, R.E. & Negri, A.P., 2008. Temperature thresholds for bacterial symbiosis with a sponge. The ISME Journal, 2 (8), 830-842.
Wedi, S.E., & Dunn, D.F., 1983. Gametogenesis and reproductive periodicity of the subtidal sea anemone Urticina lofotensis (Coelenterata: Actiniaria) in California. Biological Bulletin, Marine Biological Laboratory, Woods Hole, 165, 458-472.
Wulff, J., 1995. Effects of a hurricane on survival and orientation of large erect coral reef sponges. Coral Reefs, 14 (1), 55-61.
Wulff, J., 2006. Resistance vs recovery: morphological strategies of coral reef sponges. Functional Ecology, 20 (4), 699-708.
Zahn, R., Zahn, G., Müller, W., Kurelec, B., Rijavec, M., Batel, R. & Given, R., 1981. Assessing consequences of marine pollution by hydrocarbons using sponges as model organisms. Science of The Total Environment, 20 (2), 147-169.
Citation
This review can be cited as:
Last Updated: 30/11/2023
